Hình 1 mô tả khái quát các thành phần cấu trúc của một gene ở sinh vật nhân thực. Thành phần cấu trúc nào mang triplet sau phiên mã sẽ quy định tín hiệu mở đầu quá trình dịch mã?

"Tuyển Tập Đề Thi Tham Khảo Tốt Nghiệp THPT Năm 2025 - Sinh Học - Bộ Đề 04" là tài liệu ôn tập quan trọng dành cho học sinh lớp 12 chuẩn bị cho kỳ thi tốt nghiệp THPT năm 2025. Bộ đề được biên soạn theo cấu trúc đề thi mới nhất, bám sát chương trình học và định hướng đánh giá năng lực của Bộ Giáo dục và Đào tạo. Mỗi đề thi gồm các câu hỏi trắc nghiệm phong phú, bao quát kiến thức sinh học từ cơ bản đến nâng cao, giúp học sinh rèn luyện kỹ năng làm bài và phát triển tư duy khoa học. Đặc biệt, tài liệu đi kèm đáp án chi tiết, hỗ trợ học sinh tự kiểm tra, củng cố kiến thức và cải thiện kết quả học tập.
Câu hỏi liên quan
Ở gà, protein globin được tổng hợp ở phôi giai đoạn 14 ngày nhưng không được tổng hợp ở tế bào bạch cầu. Người ta tiến hành các thí nghiệm (TN1, TN2, TN3, TN4) theo 4 bước như ở bảng 12.
Bảng 2. Các bước thí nghiệm

Chú thích: (+) Có thực hiện bước thí nghiệm, ( – ) không thực hiện bước thí nghiệm
Kết quả cho thấy, TN1 không có các đoạn DNA có kích thước 4,6 kb (1kb = 1000 cặp nucleotide), còn các thí nghiệm TN2, TN3, TN4 đều có đoạn DNA có kích thước 4,6 kb. Biết rằng đoạn DNA chứa gene mã hóa globin được cắt bằng enzyme giới hạn BamHI có kích thước 4,6 kb, enzyme DNAase chỉ phân hủy được DNA tại vị trí không liên kết với protein.
Giải thích nào sau đây là đúng cho kết quả thí nghiệm trên?
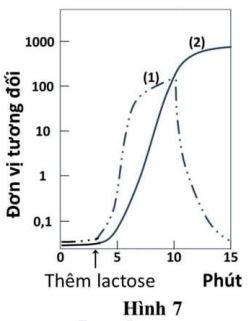
Hình 7 cho thấy kết quả xác định mức biểu hiện (qua phiên mã và dịch mã) của gene Z mã hóa enzyme \(\beta \)-galactosidase ở chủng E. coli kiểu dại, sinh trưởng trong môi trường tối thiểu ban đầu không có lactose và được bổ sung một lượng nhất định lactose vào phút thứ 3. Các đường đồ thị (1), (2) biểu thị sản phẩm của gene Z (lượng enzyme \(\beta \)-galactosidase hoặc lượng mRNA).
Phiên mã là quá trình tổng hợp nên phân tử
Bảng 4 mô tả hàm lượng mRNA và protein tương đối của gene lacZ thuộc operon lac ở các chủng vi khuẩn E. coli trong môi trường có hoặc không có lactose. Biết rằng chủng 1 là chủng bình thường, các chủng 2, 3, 4 là các chủng đột biến phát sinh từ chủng 1, mỗi chủng bị đột biến ở một vị trí duy nhất trong operon lac.
Bảng 4
Chủng vi khuẩn E.coli
Có lactose
Không có lactose
Lượng mRNA
Lượng protein
Lượng mRNA
Lượng protein
Chủng 1
100%
100%
0%
0%
Chủng 2
100%
0%
0%
0%
Chủng 3
0%
0%
0%
0%
Chủng 4
100%
100%
100%
100%
Glycine được mã hóa bởi các codon: 5’GGU3’, 5’GGX3’, 5’GGA3’, 5’GGG3’. Đây là đặc điểm nào của mã di truyền?
Trong điều hòa hoạt động gene trong operon Lac ở sinh vật nhân sơ, ngoài allolactose, trong thực nghiệm người ta cũng sử dụng một số chất để nghiên cứu về cơ chế điều hòa hoạt động gene được mô tả như Bảng 1.

Họ tiến hành bốn thí nghiệm nhằm quan sát sự tương tác của ba chất trên như sau:
- Thí nghiệm 1: môi trường có allolactose.
- Thí nghiệm 2: môi trường có chất A.
- Thí nghiệm 3: môi trường có chất A và C.
- Thí nghiệm 4: môi trường có chất B và allolactose.
Theo lí thuyết, phát biểu nào sau đây sai khi nói về thí nghiệm này?
Cho biết mạch bổ sung của gene có trình tự các đơn phân 3’…GGGAAATTT…5’. Trình tự các đơn phân tương ứng trên đoạn mạch của phân tử mRNA do gene này tổng hợp là gì?
Hình dưới đây mô tả khái quát các thành phần cấu trúc một gene ở sinh vật nhân thực. Thành phần cấu trúc nào chứa bộ ba kết thúc quá trình dịch mã?
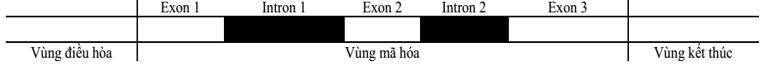
Ở vi khuẩn E. coli , biểu hiện của các gene cấu trúc trong operon lac là các gene này có số lần nhân đôi …(1)… và số lần phiên mã …(2)…
Các từ/cụm từ cần điền vào vị trí (1), (2) lần lượt là:
Polyribosome (polysome) có ý nghĩa gì trong quá trình dịch mã?
Khi không có glucose, vi khuẩn E. coli có thể sử dụng arabinose (một loại đường pentose) để làm nguồn năng lượng thay thế. Operon arabinose (hình bên) bao gồm bốn gene aramino acid, araB, araC và araD, trong đó ba gene aramino acid, araB, araD có vai trò mã hoá cho các enzyme trực tiếp tham gia vào quá trình chuyển hoá đường arabinose. Gene araC mã hoá cho một loại protein điều hoà phiên mã có khả năng liên kết với trình tự promoter của operon arabinose.
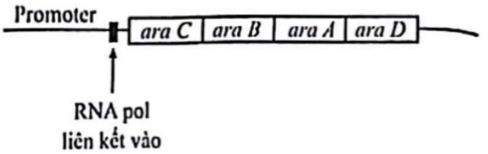
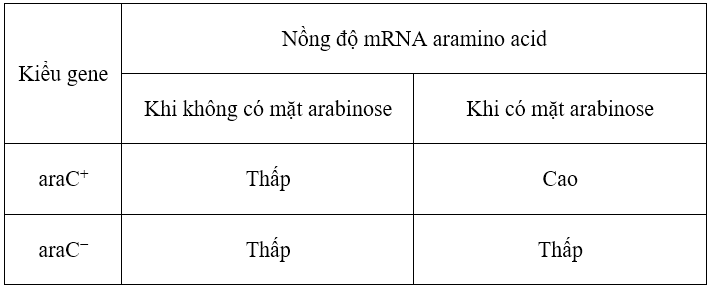
Để tìm hiểu về những đặc tính của protein AraC, các nhà khoa học đã tạo ra một chủng vi khuẩn đột biến mất chức năng gene araC và xem xét những ảnh hưởng của đột biến đó lên cường độ phiên mã của gene ara trong hai điều kiện môi trường có bổ sung hoặc không bổ sung arabinose. Những bước thí nghiệm tương tự cũng được thực hiện đối với tế bào kiểu dại để làm đối chứng. Kết quả so sánh được tóm tắt trong bảng dưới đây.
Cho đoạn mạch khuôn của gene có trình tự nucleotide như sau:
3' TAC TAT GAT TAA CAT AAT TCT GGT ATT 5'
Gene này thực hiện phiên mã và dịch mã thì số amino acid trong phân tử protein hoàn chỉnh tương ứng bằng bao nhiêu?
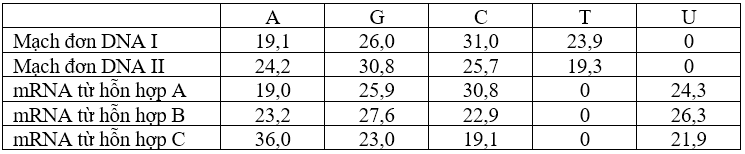
Một nhà khoa học đang nghiên cứu về trình tự của một đoạn DNA. Ông tách đoạn DNA kép này thành 2 mạch đơn rồi phân tích thành phần base Nitrogen của từng mạch. Sau khi xác định được mạch làm khuôn cho phiên mã, ông phân lập mạch này rồi bổ sung các protein cần thiết cho quá trình phiên mã xảy ra, riêng hỗn hợp B và hỗn hợp C ông còn bổ sung 1 số thành phần khác có liên quan đến xử lý mRNA. Tỉ lệ phần trăm các loại base Nitrogen của từng mạch đơn DNA và mRNA từ 3 hỗn hợp trên được thể hiện ở bảng dưới đây:
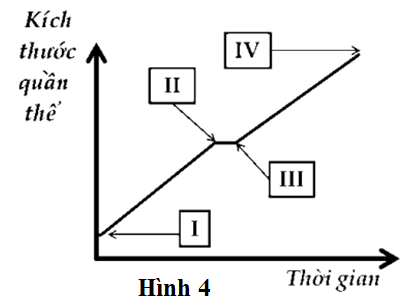
Ở vi khuẩn Escherichia coli, chức năng chính của operon Lac là giúp vi khuẩn phân giải và sử dụng được đường lactose trong tình huống môi trường chỉ có lactose là nguồn carbon duy nhất. Nếu môi trường có glucose thì operon Lac bị ức chế. Tuy nhiên, khi môi trường có cả lactose và đường glucose thì sự sinh trưởng của quần thể vi khuẩn được mô tả ở Hình 4. Theo lý thuyết, operon Lac của vi khuẩn bắt đầu hoạt động từ thời điểm nào trên đồ thị?
Vùng điều hoà của gene cấu trúc nằm ở vị trí nào của gene?
Hình 4 minh họa vai trò của protein TET trong quá trình oxygen hóa 5mC thành 5hmC và sự tương tác của yếu tố phiên mã với nucleosome để điều chỉnh biểu hiện gene. Hình 4A mô tả hoạt động của protein TET, khi TET xúc tác quá trình chuyển đổi 5-methylcytosine (5mC) thành 5-hydroxymethylcytosine (5hmC). Hình 4B minh họa vai trò của yếu tố phiên mã (TF) trong quá trình điều chỉnh hoạt động của TET. Khi TF₁ liên kết với nucleosome thì TET được gắn với vị trí tại đó, oxygen hóa 5mC thành 5hmC. TF₂ được gắn tiếp theo , với TF₁ làm thay đổi cấu trúc nhiễm sắc thể, có thể liên quan đến quá trình kích hoạt enhancer (một yếu tố tăng cường phiên mã).

Yếu tố nào có thể làm tăng hoạt động của TET?
Quá trình phiên mã ở vi khuẩn E. coli diễn ra ở
Các nhà khoa học nghiên cứu tốc độ phiên mã của một loại RNA polymerase chứa đột biến thay thế một amino acid so với chủng nấm men thông thường. Ở thí nghiệm thứ nhất, họ đã xác định được tốc độ phiên mã tối đa trong điều kiện bình thường của loại RNA polymerase trên và so sánh với RNA polymerase kiểu dại (Hình 8a).
Trong thí nghiệm thứ hai, các nhà khoa học lần lượt xử lý chủng nấm men kiểu dại và chủng nấm men đột biến với dung dịch amanitin có nồng độ 40 ng/mL và đo tốc độ phiên mã tối đa ở cả hai chủng (Hình 8b). Biết rằng hợp chất amanitin (được tìm thấy ở nhiều loài nấm độc) là một hợp chất có khả năng bám vào trung tâm hoạt động và ức chế hoạt tính của RNA polymerase.

Hình 2 mô tả các thành phần tham gia vào quá trình điều hòa hoạt động của gene trong một operon cảm ứng (operon Lac) của vi khuẩn E. coli. Sự phiên mã của các gene cấu trúc thường bị ức chế bởi một protein bám vào vùng operator (protein ức chế) ngăn cản RNA polymerase hoạt động. Khi có chất cảm ứng allolactose lactose, chất này sẽ gắn vào protein ức chế, làm thay đổi cấu trúc không gian của protein ức chế, nên protein này mất khả năng bám vào operator, cho phép RNA polymerase liên kết với promoter và bắt đầu phiên mã các gene cấu trúc.

Hình nào sau đây mô tả chính xác hoạt động của operon Lac khi môi trường có lactose nhưng các gene cấu trúc không hoạt động?

Ở vi khuẩn E. coli kiểu dại, sự biểu hiện của gene lac Z thuộc nhóm operon lac mã hóa Beta-galactoxidase phụ thuộc sự có mặt của glucose và lactose trong môi trường. Khi môi trường có cả glucose và lactose, enzyme này biểu hiện ở mức thấp, khi môi trường chỉ có lactose, enzyme được biểu hiện ở mức tăng cường trong các tế bào vi khuẩn kiểu dại. Bằng kĩ thuật gây đột biến và chuyển DNA plasmid mang các trình tự gene có nguồn gốc từ nhiễm sắc thể E. coli này vào các tế bào E. coli khác, người ta đã tạo được 3 chủng vi khuẩn đột biến có kiểu gene lưỡng bội về các gene và trình tự điều hòa tham gia phân giải lactose (chủng 1 tới 5) như Hình 7.

Trong đó:
I+, P+, O+, Z+ tương ứng là các trình tự kiểu dại của gene mã hóa protein ức chế (I) , vùng P , vùng O và gene lacZ.
P-, O-, Z- là các trình tự đột biến mất chức năng so với trình tự kiểu dại tương ứng.
I- là đột biến làm protein ức chế mất khả năng gắn vùng O.
IS là đột biến làm protein ức chế mất khả năng gắn vào đồng phân của lactose.
Xét các phát biểu dưới đây khi đưa các chủng trên vào các môi trường không có glucose và lactose, chỉ có lactose, có cả glucose và lactose.















